See how this article has been cited at scite.ai
scite shows how a scientific paper has been cited by providing the context of the citation, a classification describing whether it supports, mentions, or contrasts the cited claim, and a label indicating in which section the citation was made.
Colorimetric and fluorescent TRAP assays for visualising and quantifying fish osteoclast activity
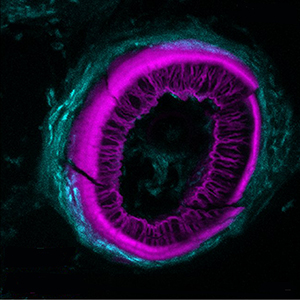
Histochemical detection of tartrate-resistant acid phosphatase (TRAP) activity is a fundamental technique for visualizing osteoclastic bone resorption and assessing osteoclast activity status in tissues. This approach has mostly employed colorimetric detection, which has limited quantification of activity in situ and co-labelling with other skeletal markers. Here we report simple colorimetric and fluorescent TRAP assays in zebrafish and medaka, two important model organisms for investigating the pathogenesis of bone disorders. We show fluorescent TRAP staining, utilising the ELF97 substrate, is a rapid, robust and stable system to visualise and quantify osteoclast activity in zebrafish, and is compatible with other fluorescence stains, transgenic lines and antibody approaches. Using this approach, we show that TRAP activity is predominantly found around the base of the zebrafish pharyngeal teeth, where osteoclast activity state appears to be heterogeneous.
Downloads
Publication Facts
Reviewer profiles N/A
Author statements
- Editor & editorial board
- profiles
- Academic society
- N/A
- Publisher
- PAGEPress Publications, Pavia, Italy

To learn about these publication facts, click ![]()
PF is maintained by the Public Knowledge Project
Citations




How to Cite

This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
PAGEPress has chosen to apply the Creative Commons Attribution NonCommercial 4.0 International License (CC BY-NC 4.0) to all manuscripts to be published.